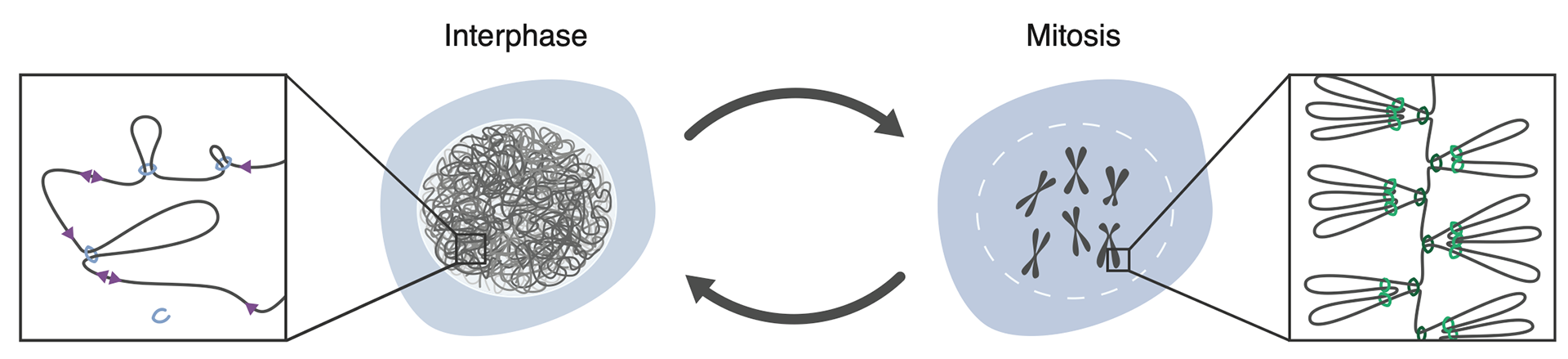
Genome organization in interphase and mitosis.
The interphase genome is organized by the formation of loops by cohesin (blue rings). Loop formation, enlargement, and release within CTCF boundaries (purple triangle) gives rise to dynamic structures. Mitotic chromosomes by contrast are compacted into rigid rod-like structures by condensin-mediated looping (green rings).
The interphase genome is organized by the formation of loops by cohesin (blue rings). Loop formation, enlargement, and release within CTCF boundaries (purple triangle) gives rise to dynamic structures. Mitotic chromosomes by contrast are compacted into rigid rod-like structures by condensin-mediated looping (green rings).
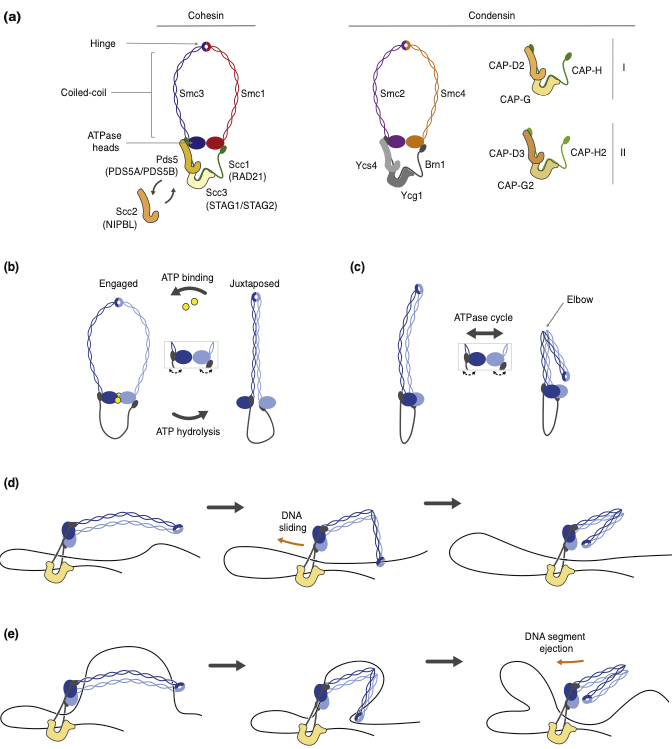
Models for how cohesin brings together CTCF sites.
(a) The genome is organized into topologically associated domains (TADs), here depicted as squares of enriched local contacts along the genome, as they are generally visualized by Hi-C experiments. These structures can be explained by the dynamic formation and enlargement of cohesin- dependent loops between CTCF boundaries. Some TADs display ‘corner peaks’ at the locations where two CTCF sites are looped together. Other TADs display a ‘stripe’ that depicts the preferential contacts of one CTCF site with sequences within the TAD. The cartoons of the loops depict an unspecified number of cohesin complexes within each blue ball.
(b) Cohesin anchoring at CTCF sites. Left: two individual cohesin complexes anchor at CTCF sites, and each then reels in DNA until the complexes collide at a random spot. Right: one individual cohesin complex anchors at a CTCF site and reels in DNA until it encounters the second CTCF site. Both scenarios lead to the looping together of two CTCF sites.
(c) Loading of cohesin dimers. Cohesin dimers, bound via their kleisin and/or regulatory subunits (yellow anchors), could reel in DNA in opposite directions. This could occur either with no anchoring to the DNA, which results in the formation of a single large loop (left), or alternatively with the use of anchors, and thus the formation of two distinct loops. In both cases, loop formation would proceed bi-directionally even if the reeling mechanism is asymmetric.
(d) Loading of multiple cohesin monomers. The loading of multiple monomers may occur in regions with a permissive chromatin context. The different monomers can reel in opposite directions. This again results in bi-directional loop formation, even though each individual complex acts in an asymmetric manner.
(a) The genome is organized into topologically associated domains (TADs), here depicted as squares of enriched local contacts along the genome, as they are generally visualized by Hi-C experiments. These structures can be explained by the dynamic formation and enlargement of cohesin- dependent loops between CTCF boundaries. Some TADs display ‘corner peaks’ at the locations where two CTCF sites are looped together. Other TADs display a ‘stripe’ that depicts the preferential contacts of one CTCF site with sequences within the TAD. The cartoons of the loops depict an unspecified number of cohesin complexes within each blue ball.
(b) Cohesin anchoring at CTCF sites. Left: two individual cohesin complexes anchor at CTCF sites, and each then reels in DNA until the complexes collide at a random spot. Right: one individual cohesin complex anchors at a CTCF site and reels in DNA until it encounters the second CTCF site. Both scenarios lead to the looping together of two CTCF sites.
(c) Loading of cohesin dimers. Cohesin dimers, bound via their kleisin and/or regulatory subunits (yellow anchors), could reel in DNA in opposite directions. This could occur either with no anchoring to the DNA, which results in the formation of a single large loop (left), or alternatively with the use of anchors, and thus the formation of two distinct loops. In both cases, loop formation would proceed bi-directionally even if the reeling mechanism is asymmetric.
(d) Loading of multiple cohesin monomers. The loading of multiple monomers may occur in regions with a permissive chromatin context. The different monomers can reel in opposite directions. This again results in bi-directional loop formation, even though each individual complex acts in an asymmetric manner.
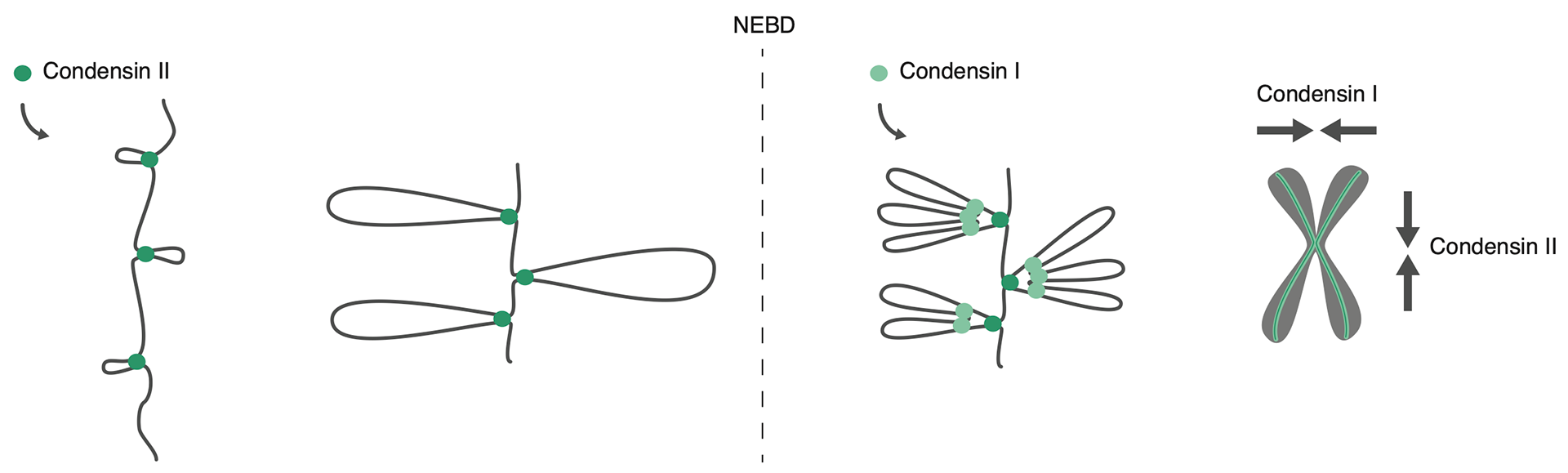
Genome organization in interphase and mitosis.
The interphase genome is organized by the formation of loops by cohesin (blue rings). Loop formation, enlargement, and release within CTCF boundaries (purple triangle) gives rise to dynamic structures. Mitotic chromosomes by contrast are compacted into rigid rod-like structures by condensin-mediated looping (green rings).
The interphase genome is organized by the formation of loops by cohesin (blue rings). Loop formation, enlargement, and release within CTCF boundaries (purple triangle) gives rise to dynamic structures. Mitotic chromosomes by contrast are compacted into rigid rod-like structures by condensin-mediated looping (green rings).
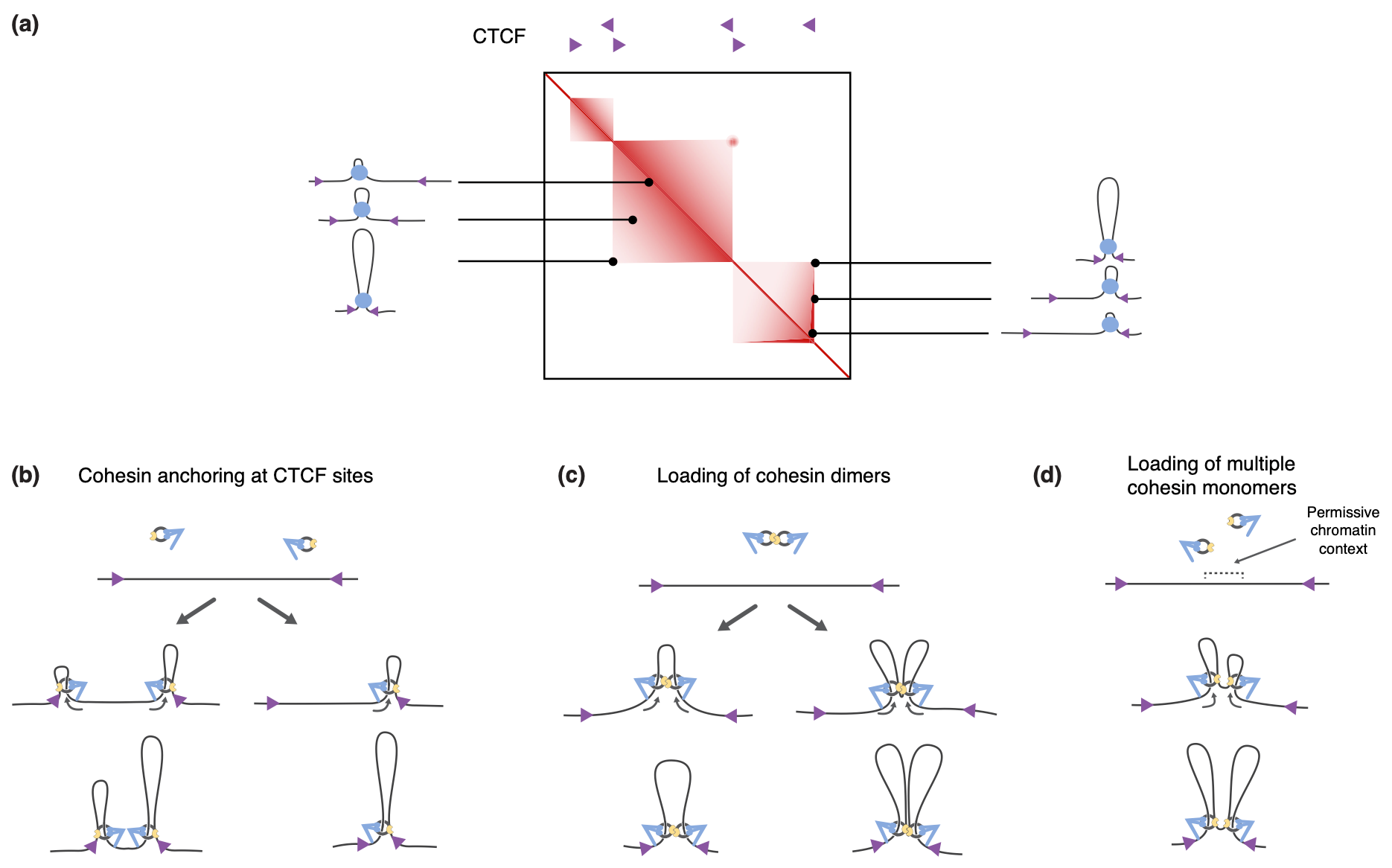
Models for how cohesin brings together CTCF sites.
(a) The genome is organized into topologically associated domains (TADs), here depicted as squares of enriched local contacts along the genome, as they are generally visualized by Hi-C experiments. These structures can be explained by the dynamic formation and enlargement of cohesin- dependent loops between CTCF boundaries. Some TADs display ‘corner peaks’ at the locations where two CTCF sites are looped together. Other TADs display a ‘stripe’ that depicts the preferential contacts of one CTCF site with sequences within the TAD. The cartoons of the loops depict an unspecified number of cohesin complexes within each blue ball.
(b) Cohesin anchoring at CTCF sites. Left: two individual cohesin complexes anchor at CTCF sites, and each then reels in DNA until the complexes collide at a random spot. Right: one individual cohesin complex anchors at a CTCF site and reels in DNA until it encounters the second CTCF site. Both scenarios lead to the looping together of two CTCF sites.
(c) Loading of cohesin dimers. Cohesin dimers, bound via their kleisin and/or regulatory subunits (yellow anchors), could reel in DNA in opposite directions. This could occur either with no anchoring to the DNA, which results in the formation of a single large loop (left), or alternatively with the use of anchors, and thus the formation of two distinct loops. In both cases, loop formation would proceed bi-directionally even if the reeling mechanism is asymmetric.
(d) Loading of multiple cohesin monomers. The loading of multiple monomers may occur in regions with a permissive chromatin context. The different monomers can reel in opposite directions. This again results in bi-directional loop formation, even though each individual complex acts in an asymmetric manner.
(a) The genome is organized into topologically associated domains (TADs), here depicted as squares of enriched local contacts along the genome, as they are generally visualized by Hi-C experiments. These structures can be explained by the dynamic formation and enlargement of cohesin- dependent loops between CTCF boundaries. Some TADs display ‘corner peaks’ at the locations where two CTCF sites are looped together. Other TADs display a ‘stripe’ that depicts the preferential contacts of one CTCF site with sequences within the TAD. The cartoons of the loops depict an unspecified number of cohesin complexes within each blue ball.
(b) Cohesin anchoring at CTCF sites. Left: two individual cohesin complexes anchor at CTCF sites, and each then reels in DNA until the complexes collide at a random spot. Right: one individual cohesin complex anchors at a CTCF site and reels in DNA until it encounters the second CTCF site. Both scenarios lead to the looping together of two CTCF sites.
(c) Loading of cohesin dimers. Cohesin dimers, bound via their kleisin and/or regulatory subunits (yellow anchors), could reel in DNA in opposite directions. This could occur either with no anchoring to the DNA, which results in the formation of a single large loop (left), or alternatively with the use of anchors, and thus the formation of two distinct loops. In both cases, loop formation would proceed bi-directionally even if the reeling mechanism is asymmetric.
(d) Loading of multiple cohesin monomers. The loading of multiple monomers may occur in regions with a permissive chromatin context. The different monomers can reel in opposite directions. This again results in bi-directional loop formation, even though each individual complex acts in an asymmetric manner.